Engineering
- Home
- Engineering
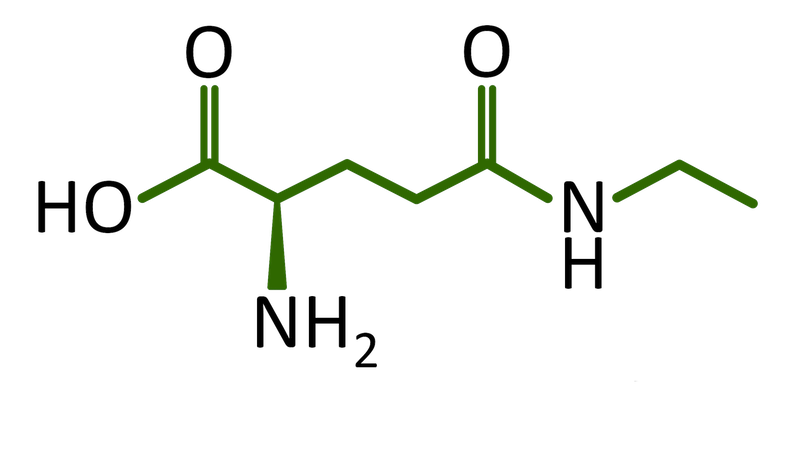
L-theanine is a non-protein amino acid found in tea leaves that can be used to alleviate anxiety and improve sleep quality. However, conventional production methods suffer from severe drawbacks. The plant extraction method is low in yield and purity, high material costs, and a risk of contamination, which can lead to significant loss of theanine. extracting theanine from tea leaves involves high costs, complex procedures, and the risk of low yields. Current chemical synthesis methods require extracting theanine from mixtures containing toxic substances, which is not only harmful to the environment but also costly.
This year, SHSBNU-China aims to use synthetic biology techniques to genetically edit the E. coli strain BL21 to produce L-theanine. Our experiment is divided into four stages. First, we designed a plasmid containing five genes (BsAld, CsAlaDC, GMAS, PPK, and GNP1) responsible for the production and transport of theanine. In the second stage, we assembled the five genes from four plasmids onto pET28a through Golden Gate Assembly, which included the design of Golden Gate primers and assembly. In the third stage, we transformed the plasmid with the five genes into the E. coli strain BL21 and induced expression with IPTG, followed by HP-LC testing of the supernatant to determine the yield. Finally, we used the yield measured by HP-LC and our human practice interviews to determine what steps we should take next to increase theanine production.
In our project, we successfully achieved a production level of 38 mg/L by genetically editing the BL21 strain. We hope to achieve higher yields in the future through batch fermentation and by improving the fermentation medium!
We used five genes in total: BsAld, CsAlaDC, GMAS, PPK, and GNP1.
- BsAld, CsAlaDC, and GMAS are responsible for the de novo synthesis of theanine, starting from glucose.
- PPK provides ATP to aid the synthesis pathway.
- GNP1 is responsible for transporting the synthesized theanine out of the bacterial cells.
In the enzymes synthesizing theanine, BsAld and CsAlaDC carry out the conversion of pyruvate to ethylamine, and GMAS synthesizes theanine from ethylamine and glutamate, which is produced by the generic bacterial TCA cycle. The coding sequence of all five enzyme were first learned from NCBI genebank, and sent for chemical synthesis by BGI genomics. The as-synthesized DNA fragments were inserted in pUC vectors. The heads and tails of each gene segment, as well as the empty vector pET28a, are designed with BsaI enzyme binding sites, allowing all gene splicing and insertion to be conducted within the same Golden Gate reaction system.

BsAld, from Bacillus subtilis, is the first step in theanine synthesis, converting pyruvate to alanine. BsAld first binds with NADH to form an enzyme-cofactor complex. Then, pyruvate binds to the active site of the enzyme-cofactor complex, and NADH transfers its electrons to the pyruvate, reducing the carbonyl group (C=O) to an imine (C=NH). The imine intermediate reacts with ammonium ions to form a Schiff base intermediate, which is then hydrolyzed to produce alanine, releasing NAD⁺ and water.
CsAlaDC, from Camellia sinensis, is the second step in theanine synthesis, converting alanine to ethylamine. CsAlaDC requires the cofactor pyridoxal phosphate (PLP), which is the active form of vitamin B6. Initially, the amino group of alanine forms a Schiff base intermediate with the aldehyde group of PLP. In the Schiff base intermediate, electrons on the α-carboxyl group are redistributed, subsequently removing α-hydrogen as carbon dioxide, forming a stable resonance structure. The resulting carbanion accepts a proton, forming the PLP-bound form of ethylamine. Ethylamine dissociates from PLP, restoring PLP to its original state, and the enzyme can proceed to the next catalytic cycle.
GMAS, from Methyloversatilis universalis, is the final step in theanine synthesis, converting ethylamine and glutamate into L-theanine, but this step requires energy supplied by ATP. Glutamate is produced by the generic bacterial TCA cycle. Therefore, this pathway is a de novo pathway, generally independent of input substrate, needing only the basic nutrients glucose for bacterial growth and intermediates synthesis.

As mentioned, the synthesis of L-theanine by GMAS requires energy from ATP. PPK, originating from E. coli, can transfer a phosphate group from the end of a polyphosphate chain to ADP, generating ATP, thereby powering GMAS. Polyphosphate is a long-chain molecule composed of multiple inorganic phosphates linked by high-energy phosphate bonds. In many bacteria and microorganisms, polyphosphate chains serve as storage forms of phosphate groups and energy.
GNP1 is responsible for transporting theanine out of E. coli. As a membrane protein, GNP1 has specific substrate binding sites. Since the structure of theanine closely resembles that of glutamine, with similar amino structures, GNP1 can recognize and bind theanine molecules. GNP1 is a proton gradient-dependent transporter, meaning that its process of transporting theanine depends on the transmembrane proton electrochemical gradient. Experimental studies have also shown that other amino acids, such as lysine and basic amino acids, cannot be transported by GNP1, indicating its strict structural requirements for specific substrates.
These five genes are located on four plasmids, and we have designed BsaI binding sites before each gene's RBS and after the stop codon. We also designed BsaI restriction sites on the pET28a expression plasmid. The table below describes the sticky ends produced by BsaI digestion of pET28a and the four gene fragments.

According to the principle of base-pair complementarity, the specificity of these fragments allows them to automatically assemble together. Ultimately, the five gene fragments were inserted into the empty vector pET28a between the lactose promoter and the T7 terminator, as shown in the following figure.


In the Golden Gate assembly, we added four pUC plasmids each containing
five genes, the empty pET28a plasmid, and the Golden Gate Kit into the
same reaction system. The content of the reaction system is as follows:
200 ng of plasmids containing GMAS, PPK, GNP1, BsAld, and CsAlaDC each,
1 μL of PGGA, 2 μL of T4 DNA ligase, 2 μL of Golden Gate Mix enzymes (which
included BsaI), and the total reaction volume was adjusted to 14 μL with
the addition of 9 μL of water. The entire Golden Gate assembly was carried
out on ice. We performed two sets of Golden Gate Assembly following the
method described above.

After the Golden Gate Assembly, we conducted DNA electrophoresis to verify whether the five gene fragments were successfully assembled. The results indicated that the Golden Gate Assembly was successful:

The verification of the success of Golden Gate assembly was performed by PCR amplification of the target connected composite coding sequence. The forward and reverse primers for the verification PCR run were design to cross two neighboring coding sequence segments and only cover about 220 bp for faster PCR cycles. Therefore, only when the coding genes are successfully connected, we can obtain a PCR product. If the assembly failed, there will be no PCR product.
Finally, we also verified the success of Golden Gate via sending the plasmid for sequencing.
After successfully obtaining the complete pET28a plasmid containing five genes via Golden Gate assembly, we transformed it into BL21 competent cells and performed plate screening. Subsequently, we divided the transformed cells into 12 tubes for fermentation in the following fermentation medium composition: 10 g/L tryptone, 5 g/L yeast extract, 10 mmol/L MgCl2·6H2O, 150 mmol/L (NaPO3)6, 15 g/L glucose. The fermentation conditions were set at 20°C and 200 rpm. We also added IPTG to eight of these tubes according to a concentration gradient of 0 mM/L, 0.25 mM/L, 0.5 mM/L, and 1 mM/L, with three tubes for each concentration gradient.
After overnight fermentation, we extracted the precipitates from two of the tubes to perform protein electrophoresis for verification. Based on the gene sequences, the estimated sizes of the proteins were approximately 39.7 kDa for BsAld, 53.6 kDa for CsAlaDC, 48.8 kDa for GMAS, 73.6 kDa for GNP1, and 80.4 kDa for PPK. Our protein electrophoresis sequencing confirmed the successful transformation of the target plasmid, and the following figure showed the successful expression of one of the enzymes, GNP1.

Following the fermentation process, we separated the supernatant and the precipitate from the remaining tubes of bacterial culture and used HPLC (High Performance Liquid Chromatography) to check if the engineered strains produced L-theanine. The HPLC results indicated that we produced L-theanine. The results are shown in the graph below:

In the HPLC data, the standard L-Theanine sample used analytical grade L-Theanine purchased from Aladdin Chemicals with greater than 98% purity. The standard sample solution was prepared at a concentration of 200mg/L. The HPLC result showed a single peak of standard L-Theanine around retention time 2.4s. The control group was prepared using the E. coli culture with the correct expression plasmid, but with no iPTG induction, nor added glucose. There is a small hump observed in the HPLC results for the control group, suggesting possible leaking expression of the designed enzymes. For the sample group, proper iPTG induction was conducted at OD600 around 0.6 – 0.8, and 15g/L glucose was added upon iPGT induction. The HPLC result demonstrated a clear rising around retention time 2.4s, indicating the production of L-Theanine.

When analyzing our experimental results, one of our key focuses is on how to enhance the production of L-theanine through a de novo biosynthetic pathway in future experiments. This could further reduce the cost and environmental impact of L-theanine production, allowing for better utilization of L-theanine in anxiety mitigation.
According to our research, one of the advantages of using a de novo synthesis method for L-theanine production is the ability to use a single strain of microorganism instead of enzymatic preparations. This approach is more robust, especially if we can significantly improve the conversion efficiency. Indeed, should we achieve a high enough enhancement in the transformation efficiency, the use of a single strain for whole-cell fermentation will likely become more cost-effective in the future.
In terms of improving conversion efficiency and yield, our analysis of the L-theanine synthetic pathway reveals that the entire process is glucose-dependent: the two precursors for L-theanine, ethylamine, and glutamate, are generated through two distinct pathways that rely on glucose input. Therefore, carbon source selection and metabolic pathway optimization will be crucial factors. Consequently, we tested various concentrations of glucose in the fermentation culture medium, ranging from 0 g/L to 20 g/L, and estimated the final L-theanine yield. Using standard L-Theanine at 0 mg/L, 50 mg/L, and 200 mg/L to produce a standard curve, our results indicate that added glucose concentrations between 15 g/L and 20 g/L could yield approximately 38 mg/L of L-theanine. The findings clearly demonstrate a strong dependence of yield on glucose supplementation levels.
Moving forward, we could consider employing both glucose and glycerol as dual carbon sources to optimize the metabolic pathway. This would allow glycerol to be independently converted into Acetyl-CoA, preventing competition during the metabolic process.
Secondly, we could delve deeper into optimizing the enzymes within the synthetic pathway by researching and testing enzymes with similar functions from different species, selecting those with the highest production yields.
Thirdly, due to time and experimental capacity constraints, our current project only attempted L-theanine production using E. coli for fermentation at the laboratory scale. For future yield enhancement, we could optimize the fermentation system further, such as considering alternative host strains (chassis) that naturally accumulate higher levels of amino acids, particularly glutamate, like actinobacteria. Additionally, we should consider experiments at larger reaction volumes for production.
Fourthly, there is room for optimization in the expression system, including aspects such as fermentation temperature, auxiliary additives (such as sodium hexametaphosphate used in this fermentation), and the pH of the reaction system.
Furthermore, another significant advantage of designing L-theanine synthesis using a de novo approach is the potential application of the produced metabolites as probiotics for anxiety mitigation, should our production method pass safety evaluations. This represents a potential application area that traditional enzymatic production methods cannot achieve.
[1] Hagihara R, Ohno S, Hayashi M, Tabata K, Endo H. Production of l-Theanine by
Escherichia coli in the Absence of Supplemental Ethylamine. Appl Environ
Microbiol. 2021 May 11;87(11):e00031-21.
https://doi.org/10.1128/AEM.00031-21
[2] Cao, R., Hu, S., Lu, Y., Wang, W., Fu, Z., & Cheng, J. (2023). Fermentative
Production of L-Theanine in Escherichia coli via the Construction of an Adenosine
Triphosphate Regeneration System. Fermentation, 9(10), 875–875.
https://doi.org/10.3390/fermentation9100875
[3] Hidese S, Ogawa S, Ota M, Ishida I, Yasukawa Z, Ozeki M, Kunugi H. Effects of
L-Theanine Administration on Stress-Related Symptoms and Cognitive Functions in
Healthy Adults: A Randomized Controlled Trial. Nutrients. 2019 Oct 3;11(10):2362.
https://www.mdpi.com/2072-6643/11/10/2362
[4] Anas Sohail A, Ortiz F, Varghese T, Fabara SP, Batth AS, Sandesara DP,
Sabir A, Khurana M, Datta S, Patel UK. The Cognitive-Enhancing Outcomes of
Caffeine and L-theanine: A Systematic Review. Cureus. 2021 Dec 30;13(12):e20828.
https://doi.org/10.7759/cureus.20828
[5] Kakuda, T., et al. (2002). "Inhibitory effect of theanine on the
binding of [3H]AMPA, [3H]kainate, and [3H]MDL 105,519 to glutamate receptors in
rat cortical neurons." Biosci Biotechnol Biochem. 66 (12): 2683-2686.
https://pubmed.ncbi.nlm.nih.gov/12596867/
[6] Nathan, P. J., et al. (2006). "The neuropharmacology of L-theanine
(N-ethyl-L-glutamine)." J Herb Pharmacother. 6 (2): 21-30.
https://pubmed.ncbi.nlm.nih.gov/17182482/
[7] Kakuda, T., et al. (2002). "Neuropsychopharmacological effects of theanine."
Nutrition. 18 (9): 843-846.
https://www.sciencedirect.com/science/article/pii/S0899900702900114
[8] Kimura, K., et al. (2007). "L-Theanine reduces psychological and physiological
stress responses." Biol Psychol. 74 (1): 39-45.
https://pubmed.ncbi.nlm.nih.gov/16930802/
[9] Wang, L., Brennan, M., Li, S., Zhao, H., Lange, K. W., Brennan, C., et al.
(2022). How does the tea L-theanine buffer stress and anxiety. Food Science and
Human Wellness, 11(1), 467-475.
https://doi.org/10.1016/j.fshw.2021.12.004
Copyright © 2024 - Content on this site is licensed under a Creative Commons Attribution 4.0 International license.
The repository used to create this website is available at gitlab.igem.org/2024/shsbnu-china.
Acknowledgements: We would like to thank our sponsors:
